Temperature and Nooksack Dace
mbakken
View all records in the stressor response library
Species Common Name
Nooksack Dace
Latin Name (Genus species)
Rhinichthys cataractae
Stressor Name
Temperature
Specific Stressor Metric
Average daily mean and maximum
Stressor Units
°C
Vital Rate (Process)
Temperature suitability; CPUE; yoy growth
Geography
Lower Fraser Valley (British Columbia)
Detailed SR Function Description
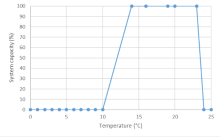
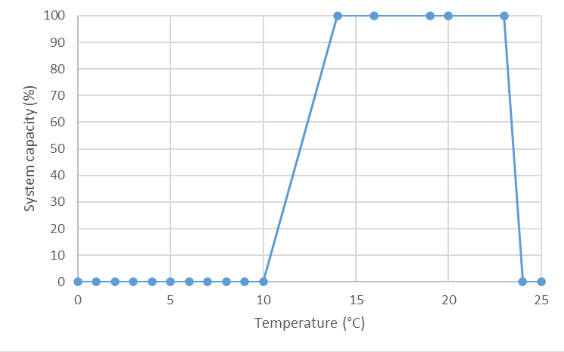
Three data sources were considered for deriving the final curve: Edwards et al. (1983) for Longnose dace temperature suitability, and Pearson (2004) and Gray and Rosenfeld (2024) for Nooksack dace CPUE and yoy growth, respectively. In addition, we considered the expert opinion provided at the workshop conducted in November 2024. Maximum system capacity was defined by the highest value for both the habitat suitability index, CPUE (fish trap-1), and fish growth, all of which were normalized to a maximum of 1 (or 100%). The overall curve (shape and threshold values) is based on the habitat suitability curve proposed by Edwards (1983) for Longnose Dace. However, the upper threshold and upper and lower y-intercepts (temperatures where system capacity is equal to zero) were modified based on existing Nooksack dace data and expert opinion. The upper threshold was first set at set at 19°C based on Edwards (1983) and then shifted to 20°C based on expert opinion (Mike Pearson, Pers. Comm., 2024) derived during a Nov. 12th 2024 expert elicitation workshop. The lower intercept value of 10°C was set based on data from Pearson (2004) for the relationship between temperature and CPUE (10°C was the lowest temperature where fish were caught in traps). The upper intercept was originally set at 23°C was based on the highest temperature where Nooksack dace were caught in traps and then shifted to 24°C based on expert opinion (Mike Pearson, Pers. Comm., 2024). Because larval dace growth experiments reported in Gray and Rosenfeld (2024) indicate higher optimal temperatures than those based on activity (CPUE) of adults, a larval/yoy dace curve was also generated with a conservatively 2C higher upper bound, assuming that individual growth effects in enclosures will be equivalent to population-level effects. Note that for general use the adult curves are most appropriate, because they are the more thermally sensitive life stage.
Maximum temperature tolerance
It should also be noted that there is anecdotal correlative evidence that Nooksack Dace are potentially more tolerant of higher temperatures (at least for short time periods) than indicated by the upper limit of our SR function. During the heat dome event of June 25- July 31, 2021, temperatures in Bertrand Creek were significantly higher than the maximum 24C that we identify as zero capacity in the temperature SR function (see flow gauge data screen shot below). The temperature sensor in the USGS Bertand Creek flow gauge at 0 Avenue on the international border recorded a high of 25.6 C on June 28th, and the accuracy of the temperature sensor is confirmed by a DO/temperature logger that was placed approximately 50 m upstream on June 30th and recorded approximately the same maximum daily temperature as the sensor at the flow gauge (data from Samantha Ramirez M.Sc. thesis; 23.3 C for USGS gauge, 23.0 C for the DO logger placed ~ 10-20cm below the surface in an unstratified glide habitat).
Although this data suggests that dace were exposed to temperatures in excess of 24C during this period, it should be noted that this exposure was of comparatively short duration - less than 12 hours on each consecutive day (see horizontal red lines in the figure below). Although groundwater inputs in lower Bertrand Creek are likely minor, it is also unclwear to what extent dace may have avoided higher temperatures by seeking out deeper water or other refuges. The impact of these thermal exposures on dace are also unclear, in terms of both potential lethal and sublethal effects. We observed dead juvenile coho and cutthroat trout in Bertrand over this time period, likely as a consequence of low dissolved oxygen and elevated temperatures. These fish were either floating or visible on the bed of the stream or stream margins. However, Nooksack dace do not have swim bladders and generally hide in substrate interstices, so that mortalities would have been unlikely to have been noticed. We did observe some unusual adult dace activity, which included observations of adults swimming during the day (adults are primarily crepuscular or nocturnal), and adult dace swimming in mixed schools of juvenile salmonids, apparently drift feeding or perhaps exploiting higher velocities to maximize oxygen exchange at the gill-water interface. However, this unusual swimming of adults in mixed schools of salmonids, although atypical, has been observed during extreme low-flow conditions when temperatures are warm in years before the Heat Dome event (Jordan Rosenfeld, pers. Obs.), and may also represent a density-dependent switch to drift feeding when benthic abundance becomes depleted. Larval (young-of-year) Nooksack dace behaviour and habitat use did not appear to be affected by the high temperatures during the heat dome based on behavioural observations and larval dace counts that were taking place during the Heat Dome event. In terms of potential population level effects, fish abundance estimates from electrofishing in Bertrand Creek riffles in Aug. 2021 (roughly 2 months after the Heat Dome) did not indicate densities of fish that were unexpectedly low, although the absence of 2020 baseline density estimates prevents a precise comparison. All in all, while the Heat Dome event does suggest some scope for tolerance of temperatures in excess of 24C for durations of up to half a day, the consequences of longer-term exposures (e.g. weeks to months) are unclear. Retaining 24C as the maximum is probably a reasonable conservative option in the short term, but simulations assessing sensitivity of cumulative effects predictions to a higher 25.6 C maximum would be informative.
Maximum temperature tolerance
It should also be noted that there is anecdotal correlative evidence that Nooksack Dace are potentially more tolerant of higher temperatures (at least for short time periods) than indicated by the upper limit of our SR function. During the heat dome event of June 25- July 31, 2021, temperatures in Bertrand Creek were significantly higher than the maximum 24C that we identify as zero capacity in the temperature SR function (see flow gauge data screen shot below). The temperature sensor in the USGS Bertand Creek flow gauge at 0 Avenue on the international border recorded a high of 25.6 C on June 28th, and the accuracy of the temperature sensor is confirmed by a DO/temperature logger that was placed approximately 50 m upstream on June 30th and recorded approximately the same maximum daily temperature as the sensor at the flow gauge (data from Samantha Ramirez M.Sc. thesis; 23.3 C for USGS gauge, 23.0 C for the DO logger placed ~ 10-20cm below the surface in an unstratified glide habitat).
Although this data suggests that dace were exposed to temperatures in excess of 24C during this period, it should be noted that this exposure was of comparatively short duration - less than 12 hours on each consecutive day (see horizontal red lines in the figure below). Although groundwater inputs in lower Bertrand Creek are likely minor, it is also unclwear to what extent dace may have avoided higher temperatures by seeking out deeper water or other refuges. The impact of these thermal exposures on dace are also unclear, in terms of both potential lethal and sublethal effects. We observed dead juvenile coho and cutthroat trout in Bertrand over this time period, likely as a consequence of low dissolved oxygen and elevated temperatures. These fish were either floating or visible on the bed of the stream or stream margins. However, Nooksack dace do not have swim bladders and generally hide in substrate interstices, so that mortalities would have been unlikely to have been noticed. We did observe some unusual adult dace activity, which included observations of adults swimming during the day (adults are primarily crepuscular or nocturnal), and adult dace swimming in mixed schools of juvenile salmonids, apparently drift feeding or perhaps exploiting higher velocities to maximize oxygen exchange at the gill-water interface. However, this unusual swimming of adults in mixed schools of salmonids, although atypical, has been observed during extreme low-flow conditions when temperatures are warm in years before the Heat Dome event (Jordan Rosenfeld, pers. Obs.), and may also represent a density-dependent switch to drift feeding when benthic abundance becomes depleted. Larval (young-of-year) Nooksack dace behaviour and habitat use did not appear to be affected by the high temperatures during the heat dome based on behavioural observations and larval dace counts that were taking place during the Heat Dome event. In terms of potential population level effects, fish abundance estimates from electrofishing in Bertrand Creek riffles in Aug. 2021 (roughly 2 months after the Heat Dome) did not indicate densities of fish that were unexpectedly low, although the absence of 2020 baseline density estimates prevents a precise comparison. All in all, while the Heat Dome event does suggest some scope for tolerance of temperatures in excess of 24C for durations of up to half a day, the consequences of longer-term exposures (e.g. weeks to months) are unclear. Retaining 24C as the maximum is probably a reasonable conservative option in the short term, but simulations assessing sensitivity of cumulative effects predictions to a higher 25.6 C maximum would be informative.
Function Derivation
Based on data from Longnose Dace, supported by data from Nooksack Dace; Empirical data (correlative model); Published; Expert opinion
Transferability of Function
This function was derived as a composite using a generic curve for Longnose Dace from across North America, and modified based on data for Nooksack Dace from Bertrand Creek and Pepin Creek - Fraser Valley. This includes observational data (catch per unit effort (CPUE) data and associated temperatures at the time minnow traps were lifted; Pearson 2004) and experimental data looking at growth of young-of-year (yoy) dace in experimental enclosures (Gray et al. 2024). As local adaptations are likely minimal among different Nooksack Dace populations. This function should therefore be broadly applicable to all populations of the species, with the caveat that it is partly based on data from other related species
Source of stressor Data
This is largely to be determined, but it will likely be a combination of data from Rosenfeld at al. (2021), data collected by Samantha Ramirez (2024) as part of her M.Sc. thesis, and a database of current and future temperature projections produced by Dr. Iacarella with DFO and collaborators (currently unpublished, but see Weller et al. 2023).
Function Type
continuous
Stressor Scale
linear
Citation(s)
Usoof, A.M. and Rosenfeld, J.S. 2024. Relationship between system capacity and spring and summer temperature for Nooksack Dace.
Edwards, E. A., H. Li, and C. B. Schreck. 1983. Habitat Suitability Index Models: Longnose Dace. U.S. Dept. Int., FishWildl. Serv. FWS/OBS-82/10.33.
Gray, J. C., and J. S. Rosenfeld. 2024. Habitat effects on recruitment, population limitations, and recovery potential in an endangered stream fish. Canadian Journal of Fisheries and Aquatic Sciences 81:1468–1480.
Pearson, M. P. 2004. The ecology, status and recovery prospects of Noonsack Dace (Rhinichthys Cataractae ssp.) and Salish Sucker (Catostomus sp.) in Canada. University of British Columbia.
Ramirez, S. Z. 2024. Modelling the impact of human development and water quality on hypoxia. University of British Columbia.
Rosenfeld, J., M. P. Pearson, J. Miners, and K. Zinn. 2021. Effects of landscape-scale hypoxia on Salish sucker and salmonid habitat associations : implications for endangered 1233:1219–1233.
Weller, J. D., R. D. D. Moore, J. C. Iacarella, J. D. Weller, R. D. D. Moore, J. C. I. Nov, and J. D. Weller. 2023. Thermalscape scenarios for British Columbia , Canada. Canadian Water Resources Journal / Revue canadienne des ressources hydriques.
Edwards, E. A., H. Li, and C. B. Schreck. 1983. Habitat Suitability Index Models: Longnose Dace. U.S. Dept. Int., FishWildl. Serv. FWS/OBS-82/10.33.
Gray, J. C., and J. S. Rosenfeld. 2024. Habitat effects on recruitment, population limitations, and recovery potential in an endangered stream fish. Canadian Journal of Fisheries and Aquatic Sciences 81:1468–1480.
Pearson, M. P. 2004. The ecology, status and recovery prospects of Noonsack Dace (Rhinichthys Cataractae ssp.) and Salish Sucker (Catostomus sp.) in Canada. University of British Columbia.
Ramirez, S. Z. 2024. Modelling the impact of human development and water quality on hypoxia. University of British Columbia.
Rosenfeld, J., M. P. Pearson, J. Miners, and K. Zinn. 2021. Effects of landscape-scale hypoxia on Salish sucker and salmonid habitat associations : implications for endangered 1233:1219–1233.
Weller, J. D., R. D. D. Moore, J. C. Iacarella, J. D. Weller, R. D. D. Moore, J. C. I. Nov, and J. D. Weller. 2023. Thermalscape scenarios for British Columbia , Canada. Canadian Water Resources Journal / Revue canadienne des ressources hydriques.
File Upload
Images
Stressor Response csv data
Data_ND_Temperature.csv
(310 bytes)
| Temperature | Mean System Capacity (%) | SD | low.limit | up.limit |
|---|---|---|---|---|
| 0 | 0 | 0 | 0 | 100 |
| 1 | 0 | 0 | 0 | 100 |
| 2 | 0 | 0 | 0 | 100 |
| 3 | 0 | 0 | 0 | 100 |
| 4 | 0 | 0 | 0 | 100 |
| 5 | 0 | 0 | 0 | 100 |
| 6 | 0 | 0 | 0 | 100 |
| 7 | 0 | 0 | 0 | 100 |
| 8 | 0 | 0 | 0 | 100 |
| 9 | 0 | 0 | 0 | 100 |
| 10 | 0 | 0 | 0 | 100 |
| 14 | 100 | 0 | 0 | 100 |
| 16 | 100 | 0 | 0 | 100 |
| 19 | 100 | 0 | 0 | 100 |
| 20 | 100 | 0 | 0 | 100 |
| 23 | 100 | 0 | 0 | 100 |
| 24 | 0 | 0 | 0 | 100 |
| 25 | 0 | 0 | 0 | 100 |
Stressor Response Chart